ISSN : 0975-5276
EISSN : 0975-9174

PRASAD M.P.1*, MANJUNATH K.2
1Sangenomics Research Labs, Bangalore- 560 071, Karnataka, India.
2Department of Microbiology, Bangalore University, Bangalore- 560 056, Karnataka, India.
* Corresponding Author : prasad_m_p@hotmail.com
Received : 30-04-2012 Accepted : 11-09-2012 Published : 29-09-2012
Volume : 4 Issue : 8 Pages : 308 - 311
Int J Microbiol Res 4.8 (2012):308-311
DOI : http://dx.doi.org/10.9735/0975-5276.4.8.308-311
The present investigation was carried out to evaluate the effect of different growth conditions on lipase production by different bacterial isolates. The extracellular lipase producing bacteria Bacillus sp., Serratia sp. Pseudomonas sp. and Staphylococcus sp. were isolated from industrial effluents. Optimization of physical and chemical parameters was done for maximum lipase production using these isolates and mixed culture (consortia). Growth of the organisms and lipase production were measured with varying pH (4-10), incubation temperature (4-60°C), incubation time (15-72 hrs.), carbon source, nitrogen source, different inducers and metal ions. Enhanced lipase production for most of the isolates was observed at 35°C, pH 7 and after 45 hrs. of incubation. Glycerol, soy meal and starch were observed as effective carbon sources and casein, peptone and tryptone as effective nitrogen sources for lipase production. Among the metal ions Ca2+ showed good lipase production with all the isolates and with consortia and the substrates Pongemia oil and Neem oil were observed as good inducers for lipase production. In the present study consortia showed an increased lipase content in comparison to individual isolates under all the optimized conditions.
Lipase, Extracellular enzyme, Optimization, Mass production, Consortia.
Lipases are serine hydrolases of considerable physiological significance and industrial potential that can catalyze numerous reactions such as hydrolysis, interesterification, esterification, alcoholysis and aminolysis [7,13] . Lipases find immense applications in food, dairy, detergent and pharmaceutical industries. Lipases are also used in oleo chemical industry and in medical applications [4] . Industrial scale extraction of lipases is carried out in bacteria, fungi, actinomycetes and cultures of plant and animal cells. Among them, microbes are metabolically versatile and hence have advantage in many industrial processes leading to the development of microbial biotechnology [2,6] . Microbial lipases are the most important group of biocatalysts used for a variety of different biotechnological applications [10,16] . Among bacteria, Achromobacter, Alcaligenes, Arthrobacter, Pseudomonas, Staphylococcus and Chromobacterium sp. have been exploited for the production of lipases. Amongst the lipase-producing organisms, Bacillus, Candida, Penicillium, Pseudomonas, Rhizomucor and Rhizopus spp. are the outstanding ones [15] . In recent years, Serratia sp. has been studied for its ability to produce lipase [1] . Pseudomonas lipases are very interesting that are not common among lipases produced by other microorganisms, such as their thermo resistance activity at alkaline pH [26] . Bacterial lipases are glycoproteins, but some extra cellular bacterial lipases are lipoproteins [10] . Considering the importance of lipase enzyme, five lipase producing bacteria Bacillus sp. 1, Bacillus sp. 2, Pseudomonas sp., Staphylococcus sp. and Serratia sp. have been isolated from different industrial effluents and different physical and chemical parameters were optimized for enhanced lipase production.
Media and chemicals for isolation and lipase assay like p-Nitrophenylpalmitate and tributyrin were of high grades and purchased from Sigma and Hi-Media Laboratories, India.
For the present study, effluent sample containing oils and fat were collected from different oil mills like groundnut oil mill, palm oil mill and dairy effluent, slaughter house waste water, soap and detergent effluent and domestic waste water in sterile bottles and plastic bags for the isolation of lipase producing organisms under laboratory condition.
For isolation of the lipase producing microorganisms, 1.0 gm. of soil/effluent sample was dissolved in 100 ml of double distilled water. It was then serially diluted (up to 10-6) and 0.5ml were plated on sterile tributyrin agar media containing 10gms/l Tributyrin, 10gms/l Tryptone, 5gms/l NaCl, 5gms/l yeast extract and 17gms/l agar. The formation of clear zone of hydrolysis around the colony on the plate indicated lipolytic microbes. Pure culture of these organisms was obtained by repeated streaking and maintained on nutrient agar slants supplemented with 1% olive oil.
The five strains used for the present study were first selected on the basis of formation of zone of hydrolysis on tributyrin agar media and indentified on the basis of morphological, biochemical and physiological characteristics according to Bergey’s manual of determinative bacteriology.
The liquid medium for optimization contained Olive oil 5%, peptone 5gm/l, yeast extract 5gm/l, glucose 5gm/l, NaCl 3gm/l and MgSO47H2O 0.5gm/l which was used as standard. Different physical and chemical parameters were altered while keeping the others constant in the media during the study for enhanced lipase production.
The effect of carbon source on lipase production was studied using fructose, lactose, sucrose, glucose, starch, mannitol, glycerol, groundnut meal and soya meal which were substituted in production medium (1%). All the carbon sources were sterilized separately and added. Organic and inorganic Nitrogen sources like peptone, soyatone, yeast extract, tryptone, beef extract, casein, ammonium chloride, ammonium nitrate, ammonium sulphate and sodium nitrate were used at (0.8% w/v) added into the media. The effect of different substrates (1% v/v ) on the production of extra cellular lipase was studied using neem oil, palm oil, pongemia oil, ground nut oil, soyabean oil, sun flower oil, olive oil, seasame oil, castor oil, hippe oil, mustard oil, coconut oil, gingly oil and cod liver oil. Metal ions used were individually added into the media for optimization. Ca2+, Mg2+, K+ Cu2+, Mn2+, Fe2+, Co2+ and Ni2+ were used in this study to check its effect on lipase production. The production medium was adjusted to different pH ranging from 4 to 10 with 0.5 variations keeping remaining media composition constant to check the effect of pH on lipase production so that the optimum pH for enhanced lipase production can be estimated. Standard lipase production media was taken and lipase production was optimized for a temperature range of 4, 15, 25, 30, 35, 40, 45, 50, 55 and 60°C to test for their effect on lipase production. The production medium was incubated under standard conditions for a time period of 15 to 70 hrs. individually on the organisms to test the effect of time on the production of lipase. The above was incubated at 37°C for 48 hrs. under shaking conditions (150 rpm).
The crude enzyme was obtained by centrifugation at 10,000 rpm, 4ºC for 10 min. Lipase activity was determined by spectrophotometric method using p-NPP (Ï-nitrophenyl palmitate) at pH 8.0 [29] . The coefficient of extinction of p-nitrophenol (p-NPP), 1.5 x 104 L/mol/cm, was determined by measuring absorbance at 410 nm after incubation for 15 min with the enzyme. One unit was defined as the amount of enzyme liberating 1µmol of Ï-nitrophenol per min at 37ºC.
Enrichment culture technique enabled the isolation of strains from oil mill effluent with lipolytic activity in tributyrin agar plate. The morphological and biochemical characterization indicated that the suspected organism were two different types of Bacillus sp.1 (SAN-L1), Bacillus sp. 2 (SAN-L15), Serratia sp. (SAN-L21), Pseudomonas sp. (SAN-L7) and Staphylococcus sp. (SAN-L11). Average pH for lipase production was observed in the range of pH 6-8 [Fig-1] . Maximum lipase activity (8.487 units/ml) was observed for consortia at pH 7.0 followed by Bacillus sp. at pH 8 (7.354 units/ml). Only the Bacillus sp. showed activity at highly alkaline pH (pH 9 and 10).
Lipase activity at different temperatures ranging from 4°C to 60°C is summarized in [Fig-2] . Maximum lipase activity (10.331 units/ml) was observed in consortia at 35°C followed by pseudomonas sp. at 35°C (8.321 units/ml).
After 45 hrs. of incubation, maximum lipase activity (10.678 Units/ml) was observed for Consortia followed by pseudomonas (8.891 Units/ml). After 70 hrs. of incubation considerable activity was still present in the Bacillus sp., Pseudomonas sp. and consortia [Fig-3] .
Lipase activity was observed for all the isolates with glycerol and starch as carbon source [Fig-4] .
Consortia showed maximum activity (11.150 units/ml) and among the isolates Pseudomonas sp. showed 9.363 units/ml activity in casein medium [Fig-5] .
Pongemia oil, neem oil and Hippe oil were good substrate for lipase production and showed higher activities in all the isolates [Fig-6] . Maximum lipase activity (10.923 units/ml) was observed with Pongemia oil in Consortia followed by Pseudomonas sp. (9.488 units/ml).
The metal ions Ca 2+, Mg²+ and K+ showed good lipase production with all the isolates [Fig-7] . Ca2+ showed maximum lipase activity (6.176 units/ml) in Consortia and Pseudomonas sp. with Fe²+ showed 6.022 units/ml activity.
We evaluated Lipase activity on the basis of p-NP as substrate for determination of lipase activity as done earlier [9] and also for characterization of Flavobacterium odoratum lipase, [5,21,24] . All the isolates showed good response between pH 6-8 including Bacillus sp. 2 which showed very good response towards pH 8. Similar results were obtained by Sharma, et al (2009) [23] . Lipase activity was highest at pH 7 for incubation period of 45 hrs. at 35°C for Staphylococcus sp. The maximum lipase activity by Pseudomonas sp. was at pH 7 at 35°C for 45 hrs. and Serratia sp. resulted lipase activity of 6.102 U/ml after 45 hour at 30°C at pH 7. Earlier, activities of bacterial lipases at chilling temperatures have been studied by Peggy, et al (2001) [14] . In our study, temperature ranges of 30-50°C gives better results which corresponds to the results obtained by Charlex L. Sanclemente and Dharam V. Vadera (1967) [3] , Mencher and Alford (1967) [11] . But, our work is carried out for the first time in lipase producing microorganisms isolated from the industrial dumping sites and also the different genus with which the work is taken up. In earlier studies lipase producing bacterial strains were isolated from coconut oil mill soil and identified as Bacillus sp. And lipase activity was maximum at pH 7.0 in 24 hrs. of incubation time. The influence of medium temperature indicated that the lipase production by the isolated strains was higher (0.001 to 0.0021 μg/ml/min) at 37°C when compared to those at 27 and 47°C [22] .
In our study optimization for incubation time revealed better lipase activity after 40-60 hrs. Maximum lipase activity was observed after 45 hrs. of incubation in pseudomonas sp. and minimum after 15 hrs. of incubation in Bacillus sp. 2. Similar reports recorded by Sangiliyandi and Gunashekharan [19] , Jane E.H. Alex and Colin [8] , Tyski, et al [28] and Sharma, et al [23] . Among the carbon sources Lipase production was good for all the isolates in glycerol and starch. Among nitrogen sources casein and peptone are good for lipase production for all the isolates. However, the maximum lipase production was recorded in Pseudomonas species in their carbon and nitrogen sources [29] . Overall lipase production of all the isolates was better in neem oil and Pongemia oil. Lipase production was maximum in pseudomonas sp. grown in Pongemia oil. However olive oil substrate treated to Pseudomonas species revealed maximum production of lipase has been stated [27] . Similar investigation was carried out by on lipase by Pseudomonas fragi [11] . Good lipase production was observed in media containing metal ions Ca²+, K+ and Fe²+. Culture grown with Ca2+ showed maximum lipase production in Bacillus sp. 1.
In all the optimization experiments maximum lipase production was obtained for consortia. So these optimization results reveal the importance of using a mixed bacterial culture for increased lipase production and its further use in industries and lipid waste management.
[1] Abdou A.M. (2003) J. Dairy Sci., 86, 127-132.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[2] Ako C., Cillard D. and Jennings B.H. (1995) J. Am. Oil Chem. Soc., 72, 1317-1321.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[3] Charles L. San Clemente and Dharam V. Vadehra (1967) Applied Microbiology, 110-113.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[4] Bornscheuer U.T., Bessler C., Srivinas R. and Krishna S.H. (2002) Trends Biotechnol., 20, 433-437.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[5] Eggert T., Pouderoyen G.V., Pencreac'h G., Douchet I., Verger R., Dijkstra B.W., Jaeger K.E. (2002) Colloids and Surfaces B: Biointerfaces., 26, 37-46.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[6] Heraldson G.G., Gudmundsson B.O., Almarsson O. (1995) J. Tetrahedron, S1, 941-952.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[7] Jaeger K.E. and Eggert T. (2002) Curr. Opin. Biotechnol., 13, 390-397.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[8] Jane E.G., Alex C. and Colin W.J. (1991) J. Gen. Microbiol.,137, 2223-2229.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[9] Labuschagne R.B., Tonder A.V. and Litthauer D. (1997) Enzyme and Microbial Technology., 21, 52-58.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[10] Macrae A.R. and Hammond A.R. (1985) Biotechnol. Genet. Eng., 3, 193-217.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[11] Mencher J.R., Alford J.A. (1967) J. Gen. Microbiol., 48, 317-328.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[12] Ota Y., Suzuki M. and Yamada K. (1968) Agr. Biol. Chem., 32, 390.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[13] Pandey A., Benjamin S., Soccol C.R., Nigam P., Krieger N. and Soccol V.T. (1999) Biotechnol. Appl. Biochem., 29, 119-131.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[14] Peggy B., Grit B. and Karsten F. (2001) Food Microbiology, 18(2), 211-215.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[15] Rapp P., Backhaus S. (1992) Enzyme Micro. Technol., 14, 938-943.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[16] Rosenau F. and Jaeger K.E. (2000) Biochimie., 82, 1023-1032.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[17] Samad M.Y.A., Razak C.N.A., Salleh A.B., Yunus W.M.Z., Ampton K. and Basri M. (1989) J. Microbiol. Methods., 9, 51-56.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[18] San Clemente C.L. and Vadehra D.V. Appl. Microbiol., 15, 110-113.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[19] Sangiliyandi G. and Gunasekaran P. (1996) Indian J. Microbiol., 36, 109.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[20] Sangiliyandi G. and Gunasekaran P. (1998) J. Microbiol. Methods, 33, 153-156.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[21] Schulte G., Bohne L., Winkler U. (1982) Can J. Microbiol., 28, 636-642.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[22] Selva Mohan T., Palavesam A. and Immanvel G. (2008) African Journal of Biotechnology, 7(15), 2728-2735.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[23] Sharma A., Bardhan D. and Patel R. (2009) Indian Journal of Biochemistry & Biophysics, 46, 178-183.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[24] Sinchaikul S., Sookkheo B., Phutrakul S., Pan F.M., Chen S.T. (2001) Protein Expr. Purif., 22, 388-398.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[25] Smith J.H. and Alford J.A. (1966) Appl. Microbiol., 14, 699.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[26] Soberon-Chavez G. and Palmeros B. (1994) Crit. Rev. Microbiol., 20, 95-105.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[27] Suzuki T., Mushiga Y., Yamane T., Shimizu S. (1988) Applied Microbiology and Biotechnology, 27, 417-422.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[28] Tyski S., Hryniewicw Z. and Jeljaszewicj Z. (1983) Biochimica. Et. Biophysica. Acta., 749, 312-317.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[29] Winkler U.K and Stuckmann M. (1979) J. Bacteriol., 138, 663-670.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[30] Xiu-Gong G., Shu-Gui C., Ke-Chang Z. (2000) Enzyme Microb. Technol., 27, 74-82.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
 | Fig. 1- Effect of pH on Lipase Activity |
 | Fig. 2- Effect of Temperature on Lipase Activity |
 | Fig. 3- Effect of Incubation Time on Lipase Activity |
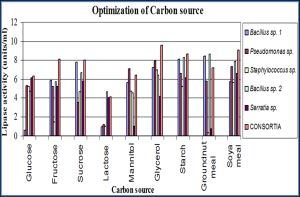 | Fig. 4- Effect of Carbon Sources on Lipase Activity |
 | Fig. 5- Effect of Nitrogen Sources on Lipase Activity |
 | Fig. 6- Effect of different Substrates on Lipase Activity |
 | Fig. 7- Effect of different Metal ions on Lipase activity |