ISSN : 0975-5276
EISSN : 0975-9174

SYAL P.1, VOHRA A.2*
1Department of Microbiology, Institute of Home Economics, University of Delhi, Hauz Khas-110 016, New Delhi, India.
2Department of Microbiology, Institute of Home Economics, University of Delhi, Hauz Khas-110 016, New Delhi, India.
* Corresponding Author : vohra_ashi@yahoo.co.in
Received : 04-03-2013 Accepted : 21-03-2013 Published : 28-03-2013
Volume : 5 Issue : 2 Pages : 390 - 398
Int J Microbiol Res 5.2 (2013):390-398
DOI : http://dx.doi.org/10.9735/0975-5276.5.2.390-398
Twenty yeast strains were isolated from traditional Indian fermented foods (idli and jalebi batter) and were screened for various probiotic properties. Seven of these isolates could survive in conditions similar to the gut with a survival rate as high as 100% at pH 2.0-2.5 and bile salt concentration of 1%. They were able to grow at 37ºC and were resistant to commonly used antibiotics. Auto-aggregation ability and cell surface hydrophobicity was observed to be high for all of the isolates. Antimicrobial action was exhibited by these isolates against enteric pathogens (i.e., E. coli, Salmonella sp., Staphylococcus aureus, Vibrio cholerae and Pseudomonas sp.). Further, they were observed to produce phytase, β-galactosidase, L-asparaginase, protease and lipase, thus could be useful in degrading anti-nutrients and improving digestion. Most of them were vitamin B12 (except J15) and exopolysaccharide producers. All of them had the ability to assimilate cholesterol in the range 57-88.5%. None of the strains produced DNase and gelatinase, thus ascertaining their safe use. These isolates were identified as Saccharomyces cerevisiae, Candida tropicalis, Aureobasidium sp. and Pichia manschuria
acid tolerance, oxbile tolerance, adhesion, safety assessment, antimicrobial, exopolysaccharide, DNase, gelatinase.
One of the oldest forms of food processing and preservation is fermentation. Three major types of microorganisms are associated with traditional fermented foods and beverages, i.e., filamentous fungi, yeasts and bacteria. Several of these non-pathogenic microorganisms, classified in the status of Generally Recognized As Safe (GRAS), are used as probiotics because of their beneficial effects on the host [1-3] .
Studies have reported the occurrence of several yeasts, such as Saccharomyces sp., Candida sp., Debaryomyces sp., Kluyveromyces sp. and Yarrowia sp., in various Indian fermented dairy and dairy-related products like curd, cheese, idli, dosa, jalebi, warries, pappad, kanji, fruit juices, bakery products and brewery products [4-6] . Several features contribute to the success of yeasts as probiotics such as their robust size, morphological diversity (budding, pseudomycelial), nutritional flexibility (ability to utilize a broad range of nitrogen, carbon, and phosphorous sources), stress tolerance ability (to low pH/oxygen/water activity, high osmotic pressure), enzyme secreting potency (secrete a broad range of enzymes such as lipase, peptidase, amylase, invertase, phytase, etc.), antioxidative/antitumor/antimicrobial activity (effective against a wide range of pathogens), and ability to produce several other useful metabolites [7] . Out of the several yeasts isolated from fermented products Saccharomyces cerevisiae according to EFSA (Europian Food Safety Authority) has a QPS (Qualified Presumption of Safety) status (Parma, Italy 2005) and is practically the only yeast commercialized as probiotic in human medicine [8,9] . It has been reported to be efficacious in the prevention or recurrence of different types of diarrhoea and colitis in humans and also has been found effective in the treatment of acute diarrhoea in children and other gastrointestinal disorders [10-12] .
The present study was undertaken with the objective of exploring the probiotic diversity of yeasts isolated from traditional Indian fermented foods (i.e, Idli and Jalebi batter). Idli, a popular traditional fermented food, prepared and consumed throughout India, is white, fermented (acid leavened) steam cake, with a soft and spongy texture, made from rice (Oryza sativum) and unhulled black gram dhal (Phaseolus mungo). Leuconostoc mesenteroides, Streptococcus faecalis and Pediococcus cerevisiae are responsible for leavening of the batter and acid production, whereas, yeasts such as Geotrichum candidum, Torulopsis lolmii, T. candida, Trichosporon pullulans, Candida sp. (C. fragilola, C. kefyr, C. tropicalis), Hansenula anomala and Rhodotorula graminis are responsible for pH reduction and may increase the thiamin and riboflavin content [13] . Jalebi, another Indian fermented food product, is a fermented sweetened product made from maida (refined wheat flour), curd and water. Lactobacillus fermentum, L. buchneri, Streptococcus lactis, S. faecalis and Saccharomyces cerevisiae are found in the fermented batter of jalebi [14] .
All of the chemicals used were of AR grade and procured from Qualigens, Himedia (Mumbai, India) and Sisco Research Laboratories (Mumbai, India).
In this investigation, idli and jalebi batter were used as the source for isolation of yeasts. The food samples were collected in sterilized reagent bottles from retail outlets in New Delhi, India and stored in an icebox till further processing.
A 25 g sample of each food sample was homogenized in 225 ml of sterile phosphate buffered saline (PBS; g L-1: NaCl 8.0, KCl 0.2, disodium phosphate 1.44 and potassium phosphate 0.24), pH 7. 10-fold serial dilutions of the samples were prepared in sterile PBS, pH 7. Malt extract-yeast extract-peptone-glucose agar (MYPG) medium (g L-1: malt extract 3.0, yeast extract 3.0, peptone 5.0, glucose 10.0 and agar 20.0, pH 5.6) was used for the isolation of yeasts. The number of yeast strains was determined by spread plating the dilutions on MYPG medium. The plates were incubated at 30ºC for 24-48 hrs. After incubation, single morphologically formed colonies were isolated, observed microscopically, sub cultured and preserved as MYPG slant agar cultures [15,16] .
Each of the yeast strain isolated was sub cultured in MYPG broth and incubated for 24-48 hrs. The cultures were centrifuged at 7000 rpm for 10 min at 4ºC. The pellets were washed and resuspended in PBS, pH 7. The effect of exposure to low pH was determined by inoculating 1% of activated yeast cultures into PBS maintained at low pH (2, 2.5, and 3) using 1N HCl. Samples were taken after 1, 3, and 5 hrs. of incubation at 37ºC. Small aliquot (0.1 mL) of the samples were plated on MYPG plates which were then incubated at 30ºC for 24-48 hrs. The survival rate was calculated as the percentage of colonies grown on MYPG medium after exposure to low pH for varying time intervals compared to the initial cell concentration [17,18] .
The effect of bile salt on the viability of the organism was monitored by inoculating 1% of yeast cultures, activated as described above, into PBS containing 0.1, 0.3, 0.5, and 1% oxbile (HiMedia), pH 7. Inoculated tubes were then incubated at 37ºC for 1, 3, and 5 hrs. and then the plate count was done at the respective time intervals. The survival rate of each strain was expressed as the percentage of viable cells in the presence of bile salt compared to that without bile salt [17,18] .
To determine the ability of the isolates to grow at human body temperature, activated yeast cultures were inoculated (106 cfu mL-1) in MYPG broth and then incubated at 30 and 37ºC for 48 hrs. The number of viable cells was determined by the plate count method and growth at both the temperatures was compared [17] .
For testing the antibiotics tolerance, the selected strains were cultivated at 30ºC for 24 hrs. The precultivated broth (0.1 mL) was spread over the MYPG medium plates. Disk diffusion method [19] , was followed for antibiotic susceptibility test with ampicillin (10 and 25 µg mL-1), chloramphenicol (30 µg mL-1), erythromycin (5 and 15 µg mL-1), penicillin G (10 µg mL-1), streptomycin (25 µg mL-1), and tetracycline (30 µg mL-1).
Auto-aggregation assay was performed according to the following method. Yeast isolates were grown for 24-48 hrs. at 37ºC in MYPG broth. The cells were harvested by centrifugation at 7000 rpm for 10 min, washed twice and resuspended in PBS, pH 7. Cell suspensions (4 mL) were mixed by vortexing for 10 sec and auto-aggregation was determined after 3 hrs. and 20 hrs. of incubation at 37ºC. An aliquot (0.1 mL) of the upper suspension of PBS after incubation was transferred to another tube with 3.9 mL of PBS and the absorbance (A) was measured at 600 nm [20] . The auto-aggregation percentage is expressed as: 1-(At/A0) X 100, where At represents the absorbance at time t = 3, or 20 hrs. and A0 the absorbance at t = 0.
For cell surface hydrophobicity, cultures of the selected strains were grown overnight at 30ºC in MYPG broth for 24-48 hrs. The cells were harvested by centrifugation at 7000 rpm, 4ºC for 10 min, washed twice and resuspended in 5 mL PBS (pH 7). Aliquots (3 mL) of the yeast suspensions were put in contact with 1 mL of each of the hydrocarbons, i.e. n-hexadecane and xylene. The cells were vortexed for 120 sec. The suspension was then kept undisturbed at 37ºC for 5 min to allow phase separation, and the hydrocarbon layer was allowed to rise completely. After 5 min, the aqueous phase was removed carefully and the absorbance (OD) was measured using a spectrophotometer at 600 nm [21] . The decrease in the absorbance was taken as a measure of the cell surface hydrophobicity (% Hydrophobicity), calculated using the equation given below:
% Hydrophobi city = (ODinitial - ODfinal) / ODinitial 100
Where, ODinitial and ODfinal are the absorbances (at 600 nm) before and after extraction with the hydrocarbons.
Spot-on-lawn method, also called double layer method, was used to evaluate the antagonistic activity of yeasts against enteric pathogens, i.e., E. coli, Salmonella sp., Staphylococcus aureus, Vibrio cholerae and Pseudomonas sp. An overnight culture of each yeast isolate was prepared in MYPG broth at 30ºC and individually inoculated onto plates by swabbing a 1 inch by 1.5-inch area in the center of each plate. The plates were incubated at 30ºC for 48 hrs. The growth in each plate was then overlaid with 10 mL of molten nutrient agar (0.7% agar) previously inoculated with 1 mL of the prepared test pathogen cultures. The agar was allowed to solidify and the plates were incubated aerobically at 37ºC for 24 hrs. and examined for growth inhibition [22,23] .
For qualitative plate assay, 100 and 20 μL of IPTG (Isopropyl β-D-1-thiogalactopyranoside, Sigma) and X-gal (5-bromo-4-chloro-indolyl-β-D-galactopyranoside, Sigma) solutions, respectively, were spread plated on the surface of PDA plates. Then the test cultures were spot inoculated on these plates and incubated at 30ºC for 24-48 hrs. Observations were then recorded as blue/white colored colonies [24] .
Phytase assay was done by point inoculating plates containing phytate minimal medium (g L-1: glucose 15.0, sodium phytate 5.0, (NH4)2SO4 5.0, MgSO4.7H2O 5.0, KCl 5.0, FeSO4.7H2O 0.01, MnSO4.4H2O 0.01 and agar 20.0) and incubating at 30ºC for 48 hrs. After incubation, the plates were flooded with 2% cobalt chloride and then kept at room temperature for 5 min. After removing cobalt chloride solution, a freshly prepared solution containing equal volume of 6.25% ammonium molybdate and 0.42% ammonium vanadate was flooded onto the plates. They were incubated at room temperature for 5 min after which, the solution was removed. The plates were observed for zone formation [25] .
A modified Czapek Dox medium (g L-1: glucose 2.0, L-asparagine 10.0, KH2PO4 1.52, KCl 0.52, MgSO4.7H2O 0.52, FeSO4.7H2O 0.01 and agar 20.0) as used for plate assay. A 2.5% stock solution of phenol red was prepared in ethanol (pH 6.2) and 3 mL of this was added to 1 L of Czapek Dox medium. Plates containing this medium were point inoculated and after 48 hrs. of incubation at 30ºC, the appearance of a pink zone around the yeast colony in an otherwise yellow medium indicates L-asparaginase activity [26] .
Casein (1%) and soluble starch (1%) were incorporated into MYPG medium to screen protease and amylase production, respectively. Plates were point inoculated and incubated at 37ºC for 48 hrs. For protease, zone formation indicates its production, whereas, for amylase production, after 48 hrs. of incubation, the plates were flooded with iodine solution and the plates were then observed for halo formation around the yeast colony against blue-black background [27,28] .
Tween 80 (1%) and phenol red (0.01%) were incorporated into MYPG medium. The pH of the medium was adjusted to 7.3-7.4 by using 1N NaOH. Inoculated plates were incubated at 37ºC for 48 hrs. and were then observed for change in color from red to yellow around the yeast colony [29] .
To determine vitamin B12 production, all of the yeast isolates were streaked on vitamin B12 assay medium (HiMedia), containing all the vitamins except vitamin B12, followed by incubation at 37ºC [30] . The growth of the isolate on the assay plate was taken as positive for vitamin B12 production.
Overnight cultures were streaked on the surface of plates containing ruthenium red milk (g L-1: skim milk powder 100.0, sucrose 10.0 and ruthenium red 0.08 and agar 20.0) [31] . After incubation at 37ºC for 24 hrs., non-ropy strains gave red colonies due to the staining of the microbial cell wall, while ropy isolates producing exopolysaccharides appeared as white colonies.
Selected yeast isolates were grown in MYPG broth, supplemented with 0.3% bile salt, at 30ºC for 24 hrs. Further, 10 mg of cholesterol dissolved in 500 µL of ethanol was added to 100 mL of MYPG broth with bile salt. The cultures were grown for 48 hrs. at 37ºC. Cells were harvested by centrifugation at 7000 rpm, 4ºC for 10 min. Supernatant was collected and used for cholesterol assay [32] . The uninoculated broth was considered as control. The percentage assimilation was calculated using the formula:
Assimilation (%) =
Gelatinase production by selected yeast isolates was studied by using tryptone-neopeptone-dextrose (TND) agar (g L-1: tryptone 17.0, neopeptone 3.0, dextrose 2.5, NaCl 5.0, K2HPO4 2.5 and agar 20.0) containing 0.4% gelatin. Petri plates containing the medium were spot inoculated with the test culture and incubated at 37ºC for 48 hrs. The petri plates were then flooded with saturated ammonium sulfate solution. Development of clear zones around the spots against the opaque background indicated a positive reaction [33] .
The selected yeast isolates were streaked on DNase agar medium (HiMedia) to check production of DNase enzyme. Plates were incubated at 30ºC for 48 hrs. After incubation a clear, pinkish zone around the colonies against dark blue background was considered positive for DNase production [33] .
The selected yeast isolates were identified by BioLog kit (IMTECH, Chandigarh, India) on the basis of phenotypic characterization.
All of the experiments were performed in triplicate. The data were assessed using analysis of variance (ANOVA) with a level of significance at p < 0.05. The results are presented as means ± standard division. All statistical analysis was performed using ‘Design Expert 6.0’ software (Stat-Ease, Inc., Minneapolis, MN, USA).
Five isolates from idli batter (i.e., Id 9, 11, 14, 15 and 18) and two isolates from jalebi batter (i.e., J 15 and 18) showed high acid tolerance at pH 2 after an exposure up to 3 hrs. with a survival rate of 56 to 100% [Table-1] Even after increasing the exposure time to 5 hrs. there was no significant decline in the survival rate. All of these isolates were also highly tolerant to bile salts [Table-2] When the concentration of oxbile was increased to 1%, there was practically no decline in the survival rate (≥ 95%).
Since, the most important prerequisite for an organism to be used as a probiotic is its ability to survive the low pH and bile in the gastrointestinal tract, therefore, out of all the yeast strains isolated, Id 9,11,14,15 and 18; J15 and 18, were screened further for more probiotic properties.
All of the strains previously selected (i.e., Id 9, Id 11, Id 14, Id 15, Id 18 and J 18) showed comparable growth at both the temperatures, i.e., 37ºC and 30ºC.
The yeast strains screened for antibiotic tolerance survived in the presence of all of the antibiotics tested as no zone of inhibition was observed around the discs.
Auto-aggregation (%) ability of the yeast isolates was determined at different time intervals. After 3 hrs. of incubation at 37ºC, except Id 9, all the isolates exhibited auto-aggregation ability above 80%. The auto-aggregation ability for all the isolates increased to 95-100% after 20 hrs. of incubation [Table-3] . These yeast isolates also exhibited 32 to 67% and 45 to 86% cell surface hydrophobicity with xylene and n-hexadecane respectively [Table-4] .
Id 15 and Id 18 strains showed antimicrobial activity against Salmonella sp., Staphylococcus aureus and Pseudomonas sp., whereas, J15 strain showed antibacterial activity against E. coli, Salmonella sp. and Vibrio cholerae. Strains Id 9, Id 11, Id 14 and J18 inhibited the growth of Pseudomonas sp. The inhibitory action was observed as a clear zone around the colonies of the yeast isolates against the lawn of the growth of pathogens. Antimicrobial activity, as measured by double layer method using different pathogenic test organisms, revealed clear circles of 16-36 mm diameter with the strains showing inhibitory action [Fig-1] .
Colonies showing blue colour production on plates containing X-Gal were considered as positive for β-galactosidase production. All of the selected strains, except J 18, were found to be β-galactosidase producers [Fig-2a] .
A clear halo in an otherwise pale yellow medium indicated phytase production, and was observed for all of the isolates, indicating them to be phytase producers [Fig-2b] .
In Czapek Dox medium, the color change in the medium from yellow to pink around the colony indicated L- asparaginase production by the isolate [Fig-2c] . Three isolates, Id 9, Id 15 and J 15, were able to produce L-asparaginase.
All of the screened yeast isolates were positive for lipase and only J 15 and J 18 strains produced protease, whereas, none of the strains showed amylase production.
All of the tested yeast isolates were observed to be positive for vitamin B12 production except J 15.
Exopolysaccharide production was observed in all of the selected yeast isolates [Fig-3] .
After 48 hrs. of incubation with cholesterol at 37ºC, yeast strains Id 9, Id 11, Id 14, Id 15, Id 18, J15 and J18 strains were found to assimilate up to 73.20%, 60.70%, 57.13%, 70.00%, 58.18%, 78.60% and 88.50% cholesterol, respectively.
None of the strains were found to be positive for gelatinase and DNase activity even after the longest incubation period, validating their relative safety as probiotic candidates.
The morphological characteristics of these seven selected isolates are listed in [Table-5] . According to BioLog kit identification, the yeast isolates were identified as Candida tropicalis (Id 9, Id 15), Saccharomyces cerevisiae (Id 11, Id 14 and Id 18), Aureobasidium sp. (J 15) and Pichia manschurica (J 18).
Yeasts are used in food fermentations since the ancient times. Several yeast strains are present in dairy related products or occur as a part of gastrointestinal tract. However, their use as probiotic agents is limited only to strains of the genus Saccharomyces. In this study, twenty yeast strains were isolated from idli and jalebi batter, which were then purified. Further, in vitro screening of these strains was done for probiotic attributes.
Probiotics need to survive the inevitable biological barriers of gut. The primary barrier of microorganisms in the stomach is the gastric acidity (pH of 1.5-3.5). Besides the strong acid condition in the stomach, the probiotic microorganisms taken orally have to defend against the bile salt in the gastrointestinal tract. Hence, bile tolerance is also considered to be one of the important properties required for high survival of the probiotic organism. There is no consensus about the precise concentration to which the selected strains should be tolerant. The physiological concentration of bile salts in the small intestine is between 0.2 and 2.0% [34] . In this investigation, Id 9,11,14,15 and 18; J15 and 18 exhibited high acid and bile salt tolerance. Similar results have been reported by several in vitro studies which indicate that yeasts belonging to Saccharomyces, Debaryomyces, and Kluyveromyces species are extremely tolerant low pH and high bile salt concentration (upto 1% oxgall) [1,2,5,24,35] .
A probiotic organism must also be able to tolerate and grow at human body temperature of 37ºC and so were these selected seven isolates. Similarly, in an investigation [36] , yeast strains isolated from chicken feces and kefir were able to grow at 37ºC. It has also been reported that the growth of Saccharomyces cerevisiae at 37ºC is a variable characteristic [17] .
Antibiotics widely used include streptomycin, tetracycline, erythromycin, chloramphenicol, penicillin and ampicillin [37] . Most of the probiotic microorganisms are bacteria and many of them are not able to resist or tolerate these antibiotics, whereas, yeasts have a natural resistance against these antibiotics and can be used for patients undergoing antibiotic treatment [18] .
Apart from tolerating the upper gastrointestinal transit, another challenge for an effective probiotic is to adhere to small intestinal cells. Adhesion to intestinal epithelial cells is an important prerequisite for colonization of probiotic strains in the gastrointestinal tract, preventing their immediate elimination by peristalsis and providing a competitive advantage in this ecosystem [38] . Adhesion is initially based on non-specific physical interactions between two surfaces, which then enable specific interactions between adhesins (usually proteins) and complementary receptors. Auto-aggregation of probiotic strains is necessary for adhesion to intestinal epithelial cells [39] . It has been reported that auto aggregation ability above 80% is considered to be strong auto-aggregation and S. cerevisiae strains with 85% auto aggregation have been reported [40] . Another important property that has been implicated in the attachment of the microorganism to the intestinal epithelial cells is the hydrophobic nature of the outermost surface of microorganisms [41] . Strains possessing high hydrophobicity exhibit good adhesion property to intestinal cell lines [42] . In general, the hydrophobicity values for indigenous yeast isolates are overall lower than the values as high as 79.69% reported for bacteria [43] . In an experiment, adhesion attributes of S.cerevisiae strains such as cell surface hydrophobicity (58.21 ± 1.09%) and auto-aggregation (67.42 ± 0.33%) have been reported [25,44] . Whereas, in this investigation, all of the yeast isolates showed high auto-aggregation ability, which was observed to be upto 94% after 3 hrs. of incubation at 37°C and after 20 hrs. it was observed to be higher than 95% for all the isolates. The cell surface hydrophobicity of 86.14%, observed for J 18 in this investigation, is higher than the hydrophobicity reported till now. It is also evident from [Table-3] and [Table-4] that there is a difference in behavior of yeast isolates for different hydrocarbons. It has been reported that yeast strains usually show variable adherence property and need to be administered repeatedly to achieve steady-state concentrations in the colon [5] . The variation in hydrophobicity to solvents has been explained by the fact that adhesion depends upon the origin of the strains as well as their surface properties [45] . Isolates having good auto-aggregation ability in conjunction with the good hydrophobicity values can strongly be related to the adhesion ability of these microorganisms. Though these traits are independent of each other; they are still related to adhesion property of a particular microbe [46] .
One of the most desirable properties of probiotic yeasts is the antibacterial activity of yeasts against pathogens that penetrate various mucosa sites [47] . Therefore, the effect of the isolated yeast strains on the growth of selected food borne pathogens was determined and antimicrobial activity was observed against E. coli, Staphylococcus aureus, Pseudomonas sp., Vibrio sp. and Salmonella sp. Several studies using animal models or cell models have indicated that S. boulardii may exert a beneficial effect against various enteric pathogens such as Clostridium difficile, Vibrio cholerae, Salmonella, Shigella and E. coli and several other yeast strains have also been shown to have antibacterial activity [48,49] . In an investigation probiotic yeast S.cerevisiae var boulardii caused a statistically significant reduction in the number of cells of Listeria monocytogenes, Pseudomonas aerugenosa and Staphylococcus aureus and a decrease in the growth rate of Listeria monocytogenes and Pseudomonas aerugenosa [50] . Yeasts act against enteric pathogens by many different mechanisms, such as prevention of bacterial adherence and translocation in the intestinal epithelial cells, production of factors that neutralize bacterial toxins and modulation of the host signaling pathway with proinflammatory response during bacterial infection [8,51,52] .
An important way in which probiotics beneficially affect the health of the host is by providing enzymatic activities that improve the utilization of nutrients within the intestine [53] . In a study, the extacellular enzymatic profiles of yeasts isolated from the tropical environments of Brazilian rain forest were explored and observed to be a source of industrially relevant enzymes such as amylases, esterases, lipases, proteases, pectinases, and chitinases [54] . Candida famata isolated from pure oil waste expressed several enzymatic activities, i.e., phosphatase, esterase, lipase and amylase [55] . The intraintestinal hydrolysis of lactose by β - galactosidase (EC 3.2.1.23) enzyme is considered as the primary mechanism in inducing a positive impact on the host’s health as this enzyme helps in avoiding problems of lactose intolerance in individuals due to deficiency of the enzyme and was observed for most of selected yeast isolates in this study [43,56] . Yeast species belonging to the genus Kluyveromyces (such as K. lactis, K. marxianus) have been used commercially for β-galactosidase production [57] . Phytases (myo-inositol-hexakisphosphate 6-phosphohydrolases) are nonspecific phosphatase enzymes, which catalyze the dephosphorylation of phytate and release free inorganic phosphate (Pi) and inositol phosphate esters [58] Phytic acid or phytate (myo-inositol hexakisphosphate, IP6) is the primary storage form of phosphorus in mature seeds of plants and has antinutritional properties since it has a strong chelating capacity and lowers the bioavailability of divalent ions such as iron, zinc, calcium and magnesium by forming insoluble complexes and also can negatively influence the functional and nutritional properties of proteins such as digesting enzymes [59,60] . In this investigation, allof the seven isolates were phytase producers. A number of yeasts are known to produce phytase such as Candida krusei, Schwanniomyces castellii, Debaryomyces castellii, Arxula adeninivorans, Pichia sp., Cryptococcus laurentii, Rhodotorula gracilis and S. cerevisiae [25,61,62] etc. Two probiotic yeasts isolated from sea cucumber Holothuria scabra namely, Yarrowia lypolytica and Candida tropicalis, have been reported to produce phytase [63] . Present market trends clearly indicate that there is an increasing demand for phyase as a feed supplement to improve feed intake and utilization. The enzyme L-asparaginase (E.C.3.5.1.1) is used as a therapeutic agent in the treatment of acute lymphoblastic leukaemia, that depletes tumor cells of L-asparagine and the cells die because of their inability to synthesize this amino acid [64] . Microorganisms producing L-asparaginase are either bacteria or fungi; very few types of yeast have been reported to produce this enzyme and was produced by Id 9, Id 15 and J 15 strains in this study.
Yeasts are a source of vitamins, especially B-complex including B11 (thiamine), B2 (riboflavin), B3 (niacin), B5 (pantothenic acid), B6 (pyridoxine), B9 (folic acid), and H or B7 (biotin). These vitamins play an important role in various metabolic processes in the body such as in breaking down carbohydrates, fats, and proteins and provide the body with energy. They also support the nervous system; help maintain the muscles used for digestion [65] and all of the tested yeast isolates were observed to be positive for Vit B12 production.
Exopolysaccharides (EPSs) are exocellular polymers and are thought to play a role in protection against desiccation, toxic compounds, bacteriophages, osmotic stress, and to permit adhesion to solid surfaces and biofilm formation [66] Another physiological benefit is that EPS is retained longer in the gastrointestinal tract, so that colonization by the probiotic microorganisms can be enhanced [67] . Consequently, EPS producing probiotic cultures can contribute human health by positively affecting the gut microflora [68] . S. cerevisiae strains, isolated traditional fermented food products, producing exopolysaccharides have been previously reported [43] . EPS has immunostimulatory [69] and antitumoral activities [70] , and phosphate groups in EPS play an important role in the activation of macrophages and lymphocytes [71] . Therefore, the yeast isolates were screened for the production of EPS and all of the strains tested positive for EPS production and can be used as immune-stimulatory adjuvants.
High levels of serum cholesterol have been associated with the risk of coronary heart disease and also in inducing colon cancer [72] . Due to the emergence of a more health-conscious society the role of biological procedures (probiotic food products) has attracted much attention. In this study, all of the yeast strains were found to remove cholesterol, with the J18 strain showing the highest potential to assimilate cholesterol. Several studies have shown that the cholesterol-lowering action of probiotics may be achieved through cholesterol assimilation [73] . S. boulardii, P. kudriavzevii and S. cerevisiae have previously been evaluated as potential probiotics for assimilating cholesterol over the past few years [74] . As a feed additive, probiotic yeast S.cerevisiae var. boulardii has been reported to reduce serum cholesterol in animals [75,76] . It has been recorded that even 1% reduction in cholesterol can reduce the risk of cardiovascular diseases by 2-3% [77] , so the results of this investigation can be considered as promising.
New species and more specific strains that are potential probiotic candidates are constantly being identified and are gradually increasing and these cannot be assumed to share the same historical safety of tested or traditional strains [78] . According to FAO/WHO (2002) every probiotic strain needs to be assessed for safety to be used as a food or feed supplement [79] . Gelatinase and DNase are the enzymes produced by mostly pathogenic microorganisms. Gelatinases (MMP-2 and MMP-9) are MMPs capable of degrading almost all ECM and basement membrane components and might provide suitable substrate for further activity of human gelatinases or other bacterial proteinases [80] . Extracellular DNase provides a growth advantage to the pathogen by enlarging the pool of available nucleotides by DNA hydrolysis helps in the dissemination and spread of the pathogen by liquifying pus and also aids the evasion of the innate immune response by degrading neutrophil extracellular traps (NETs) [81] . Some yeasts and yeast like pathogenic microorganisms, such as Cryptococcus sp., Endomycopsis sp., Tremella and Rhodotorula sp., have been known to produce extracellular DNase [82] . A microorganism should not produce these enzymes so as to be used as a probiotic in food and feed and none of the strains in this study produced DNase or gelatinase.
Saccharomyces cerevisiae is the only clinically proven yeast being used as a human probiotic and has shown to positively influence host’s health by antimicrobial effect, nutritional effect, inactivation of bacterial toxins, quorum sensing, trophic effects, immuno-modulatory effects, anti-inflammatory effects, cell restitution and maintenance of epithelial barrier integrity [83,84] . Candida tropicalis is a well-known yeast species found all over the world, and not only colonizes animal’s gut has been known to exert nutritional and other nutritional probiotic effects in the host, such as production of phytase [85] . Pichia sp. has also been known to have beneficial effects like biodegradation of phytate and folate biofortification [6,59] . Aureobasidium sp., especially, Aureobasidium pullulans, popularly known as black yeast, ubiquitous, polymorphic and oligotrophic yeast like microfungus occurs frequently in wide range of tropical and temperate environment with fluctuating moisture content in phyllospere, and also isolated from damp indoor surfaces, food and feed substances [86] . It produces a number of by-products including several enzymes (such as amylase, protease, lipase, xylanase and cellulose), has high protein content allowing it to be used as single cell protein and produces an important polysaccharide namely pullulan [87] . It has not been used as a probiotic till now, but since it has been reported to have many beneficial attributes and in this investigation also it has been observed that it can produce phytase, β-galactosidase, exopolysaccharides and has antimicrobial activity against enteric pathogens, it can serve as a potential probiotic in future.
Fermented foods are a source of diverse microflora and the fact that very few types of yeast have been commercially used as probiotic, it is worthwhile to explore the diversity of yeasts in traditional Indian fermented foods and study their probiotic attributes. Through this research we have isolated yeasts with potential probiotic properties. They could be prescribed to patients on prolonged antibiotic treatment, protect against food borne enteric pathogens. They have the ability to degrade anti-nutrients such as phytic acid and tannic acid, thereby, improving the nutritional status of food. They can hydrolyze lactose, thus prove beneficial to lactose intolerant patients. They also produce L-asparaginase which has anticancer property. These yeast isolates could aid in digestion because of their ability to produce lipase and protease. Enhance vitamin B12 pool due to its production by these isolates. They can be used as immune-stimulants. Their ability to assimilate cholesterol could be of immense value to patients suffering from hypercholesterolemia. Beneficial attributes of Saccharomyces cerevisiae, Candida sp., Pichia sp., and Aureobasidium sp. indicates them to be promising probiotic agents in future and can be widely used as food and feed supplements.
We would like to thank the Department of Science and Technology, New Delhi, India for financial assistance for this investigation.
[1] Kourelis A., Kotzamanidis C., Litopoulou-Tzanetaki E., Scouras Z.G., Tzanetakis N., Yiangou M. (2010) Journal of Biological Research-Thessaloniki, 13, 93-10.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[2] Kourelis A., Zinono I., Kakagianni M., Christidou A., Christoglou N., Yiannaki E., Testa T., Kotzamanidis C., Litopoulou-Tzanetaki E., Tzanetakis N., Yiangou M. (2010) Journal of Applied Microbiology, 108, 274-284.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[3] Tamang J.P. (1998) Indian Food Industry, 17, 162-166.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[4] Mahasneh A.M., Abbas M.M. (2010) Journal of Biological Sciences, 3, 133-140.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[5] Kumura H., Tanoue Y., Tsukahara M., Tanaka T., Shimazaki K. (2004) Journal of Dairy Science, 87, 4050-4056.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[6] Moslehi-Jenabian S., Lindegaard Pedersen L., Jespersen L. (2010) Nutrition, 2, 449-473.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[7] Fredlund E., Druvefors U., Boysen E.M., Lingsten K., Schnurer J. (2002) FEMS Yeast Research, 2, 395-402.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[8] Czerucka D., Piche T., Rampal P. (2007) Alimentary Pharmacology and Therapeutics, 26, 767-778.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[9] Etienne-Mesmin L., Livrelli V., Privat M., Denis S., Cardot J., Alric M., Blanquet-Diot S. (2011) Applied Environmental Microbiology, 77, 1127-1131.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[10] Cetina S.G., Siemo B.G. (1999) Annales de Pediatrie, 41, 397-400.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[11] McFarland L.V. (2010) World Journal of Gastroenterology, 16, 2202-2222.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[12] Kelesidis T. (2012) Therapeutic Advances in Gastroenterology, 5, 111-125.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[13] Sridevi J., Halami P., Vijayendra S. (2010) Journal of Food Science and Technology, 47, 557-563.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[14] Steinkraus K.H. (1996) Handbook of Indigenous fermented foods, New York.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[15] Erdogrul O., Erbilir F. (2006) Turkish Journal of Biology, 30, 39-44.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[16] Corbaci C., Ucar F.B., Yalcin H.T. (2012) African Journal of Microbiology Research, 6, 534-542.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[17] Barnett J.A., Payne R.W. and Yarrow D. (2000) Yeasts: Characteristics and Identification, 3rd ed., Cambridge Univ. Press, Cambridge, UK.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[18] Nayak S.K. (2011) Probiotics, Microbiology Monographs, 21, 29-55.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[19] Bauer A.W., Kirby W.M.M., Sherris J.C., Turck M. (1996) American Journal of Clinical Pathology, 45, 493-496.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[20] Del Re B., Sgorbati B., Miglioli M., Palenzona D. (2000) Letters in Applied Microbiology, 31, 438-442.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[21] Rosenberg M., Gutnick D., Rosenburg E. (1980) FEMS Microbiology Letters, 9, 29-33.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[22] Tagg J.R., McGiven A.R. (1971) Bacteriological Reviews, 40, 722-756.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[23] Maia O.B., Duarte R., Silva A.M. (2001) Veterinary Microbiology, 79, 183-189.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[24] Sourabh A., Kanwar S.S., Sharma O.P. (2011) Journal of Yeast Fungal Research, 2, 17-126.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[25] Vohra A., Satyanarayana T. (2001) Biotechnology Letters, 23, 551-554.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[26] Gulati R., Saxena R.K., Gupta R. (1997) Letters in Applied Microbiology, 24, 23-26.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[27] Montville T.J. (1983) Applied Environmental Microbiology, 45, 200-204.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[28] Selvam K., Vishnupriya B., Bose V.S.C. (2011) International Journal of Pharmaceutical and Biological Archive, 2, 1481-1487.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[29] Samad M.Y.A., Razak C.N.A., Salleh A.B., Yunus W.M.Z.W., Ampon K., Basri M. (1989) Journal of Microbiological Methods, 9, 51-56.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[30] Bishnoi K., Kataria M., Singhal V., Gupta D. (2012) International Research Journal of Pharmacy, 3, 74-82.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[31] Mora D., Fortina M.G., Parini C., Ricci G., Gatti M., Giraffa G., Manachini P.L. (2002) Journal of Applied Microbiology, 93, 278-287.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[32] Searcy R.L., Bergquist L.M. (1960) Clinica Chimica Acta., 5, 192-199.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[33] Gupta H., Malik R.K. (2007) Lait, 87, 587-601.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[34] Gunn J.S. (2000) Microbes and Infection, 2, 907-913.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[35] Pardo S., Galvagno M.A., Cerrutti P. (2009) Revista Iberoamericana de Micologia, 26, 155-160.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[36] Rajkowska K., Kunicka-Styczyńska A. (2010) Polish Journal of Microbiology, 59, 257-263.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[37] Kim S., Kim H., Chae H.J. (2004) Agricultural Chemistry and Biotechnology, 47, 20-26.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[38] Kos B., Suskovic J., Vukovic S., Simpraga M., Frece J., Matosic S. (2003) Journal of Applied Microbiology, 94, 981-987.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[39] Perez P.F., Minnaard Y., Disalvo E.A., de Antoni G.L. (1998) Applied and Environmental Microbiology, 64, 21-26.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[40] Pizzolitto R.P., Armando M.R., Combina M., Cavaglieri L.R., Dalcero A.M., Salvano M.A. (2012) Journal of Environmental Science and Health, Part B: Pesticides, Food Contaminants and Agricultural Wastes, 47, 933-941.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[41] Schillinger U., Guigas C., Holzapfel W.H. (2005) International Dairy Journal, 15, 289-1297.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[42] Pan W.H., Li P.L., Liu Z. (2006) Anaerobe, 12, 148-152.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[43] Collado M.C., Meriluoto J., Salminen S. (2008) European Food Research Technology, 226, 1065-1073.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[44] Sourabh A., Kanwar S.S., Sharma O.P. (2012) African Journal of Biotechnology, 52, 11447-11454.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[45] Morata De Ambrosini V., Gonzalez S., De Ruiz Holgando A.P., Oliver G. (1998) Journal of Food Protection, 61, 557-562.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[46] Rahman M.M., Kim W.S., Kumura H., Shimazaki K. (2008) World Journal Microbiology and Biotechnology, 24, 1593-1598.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[47] Boirivant M., Strober W. (2007) Current Opinion in Gastroenterology, 23, 679-692.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[48] Czerucka D., Rampal P. (2002) Microbes and Infection, 4, 733-739.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[49] Nikolaou E., Soufleros E.H., Bouloumpasi E., Tzanetakis N. (2006) Food Microbiology, 23, 205-211.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[50] Rajkowska K., Kunicka-Styczynska A., Rygata (2012) Food Technology and Biotechnology, 50, 230-236.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[51] Martins F.S., Elian S.D.A., Vieira A.T., Tiago F.C.P., Martins A.K.S., Silva F.C.P. (2011) International Journal of Medical Microbiology, 301, 359-364.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[52] Tiago F.C.P., Martins F.S., Souza E.L.S., Pimenta P.F.P., Araujo H.R.C., Castro I.M., Brandao R.L., Jacques R. (2012) Journal of Medical Microbiology, doi:10.1099/jmm.0.042283-0.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[53] Mustapha A., Jiang T., Savaiano D. (1997) Journal of Dairy Science, 80, 537-1545.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[54] Buzzini P., Martini A. (2002) Journal of Applied Microbiology, 93, 1020-1025.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[55] Mahdhi A., Hmila Z., Behi A., Bakhrouf A. (2011) Iranian Journal of Microbiology, 3, 129-134.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[56] Rajakala P., Selvi P.K. (2006) International Journal of Dairy Science, 1, 167-172.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[57] Dagbagli S. (2008) Electronic Journal of Biotechnology, 11, 1-12.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[58] Vucenik I., Shamsuddin A.M. (2006) Nutrition and Cancer, 55, 109-125.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[59] Vohra A., Satyanarayana T. (2003) Critical Reviews in Biotechnology, 23, 29-60.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[60] Reddy N.R., Pierson M.D. (1994) Food Research International, 27, 281-290.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[61] Lim M.H., Lee O.H., Chin J.E., Ko H.M., Kim I.C., Lee H.B., Im S.Y., Bai S. (2008) Biotechnology Letters, 30, 2125-2130.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[62] Olstorpe M., Schnurer J., Passoth V. (2009) FEMS Yeast Research, 9, 478-488.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[63] Hirimuthugoda N.Y., Chi Z. and Longfei Wu. (2007) SPC Beche de Mer Informat Bulletin 26.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[64] Verma N., Kumar K., Kaur G., Anand S. (2007) Critical Reviews in Biotechnology, 27, 45-62.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[65] Abramov S.A., Kotenko S.T., Ramazanov A.S., Islamova F.I. (2003) Prikladnaia Biokhimiia Mikrobiologiia, 39, 438-40.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[66] Vuyst L., Degeest B. (1999) FEMS Microbiology Reviews, 23, 153-177.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[67] Looijesteijn P., Trapet L., Vries E., Abee T., Hugenholtz J. (2001) International Journal of Food Microbiology, 64, 71-80.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[68] Darilmaz D.O., Aslım B., Suludere Z., Akca G. (2011) Brazilian Archives of Biology and Technology, 54, 917-926.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[69] Hosono J., Lee A., Amenati M., Natsume M., Hirayama T., Adachi S., Kaminogawa S. (1997) Bioscience, Biotechnology and Biochemistry, 61, 312-316.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[70] Ebina T., Ogata N., Murata K. (1995) Biotherapy, 9, 65-70.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[71] Uemura J.N., Kitazawa H., Kawai Y., Itoh T., Oda M., Saito T. (2003) Food Microbiology, 20, 267-273.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[72] Reddy B.S., Mastromarini A., Wynder E. (1977) Cancer, 39, 1815-1819.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[73] Dilmi-Bouras A. (2006) Annals of Agricultural and Environmental Medicine, 13, 49-53.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[74] Psomas E.I., Fletouris D.J., Litopoulou-Tzanetaki E., Tzanetakis N. (2003) Journal of Dairy Science, 86, 3416-3422.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[75] Paryad A., Mahmoudi M. (2008) African Journal of Agricultural Research, 3, 835-842.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[76] Hassanein S.M., Soliman N.K. (2010) Journal of American Science, 6, 159-169.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[77] Manson J.E., Tosteson H., Ridker P.M., Satterfield S., Hebert P., O’Connor G.T., Buring J.E., Hennekens C.H. (1992) The New England Journal of Medicine, 326, 1406-1416.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[78] Salminen S.J., von Wright A.J., Ouwehand A.C. and Holzapfel W.H. (2001) Fermentation and Food, Gaithersburg, Aspen Publishers, 239–251.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[79] FAO/WHO (2002) Guidelines for the evaluation of probiotics in food. Food and Agriculture Organization of the United Nations and World Health Organization Working Group Report, London Ontario, Canada.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[80] Zhao Q., Liu Z., Xue Y., Wang J., Li H., Tang Q., Wang Y., Dong P., Xue C., Ds-echinoside A. (2011) Journal of Zhejiang University Science B, 12, 534-544.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[81] Hasegawa T., Minami M., Okamoto A., Tatsuno I., Isaka M., Ohta M. (2010) Microbiology, 186, 184-190.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[82] Sanchez M., Colom F. (2010) Revista Iberoamericana de Micologia, 27, 10-13.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[83] Suharja A.A.S., Henriksson A., Liu S.Q. (2012) Journal of Food Processing and Preservation, doi:10.1111/j.1745-4549.2012.00 780.x.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[84] Lukaszewics M. (2012) Probiotics, 385-398, DOI: 10.5772/501 05.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[85] Chi Z.M., Liu G., Zhao S., Li J., Peng Y. (2010) Applied Microbiology and Biotechnology, 86, 1227-1241.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[86] Samson R.A., Hoekstra E.S., Frisvad J.C. (2004) Centraalbureau voor Schimmelcult, Utrecht, 7, 150-177.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[87] Gaur R., Singh R., Gupta M., Gaur M.K. (2010) African Journal of Biotechnology, 9, 7989-7997.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
 | Fig. 1- Plates showing zone of growth inhibition against lawn of bacterial growth. |
 | Fig. 2a- Plate showing β-Galactosidase enzyme production |
 | Fig. 2b- Plate showing Phytase enzyme production |
 | Fig. 2c- Plate showing L-asparaginase enzyme production |
 | Fig. 3- Exopolysaccharide production by yeast isolates |
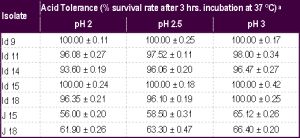 | Table 1- Survival rate of the isolates (Id 9, 11, 14, 15 and 18 from idli batter; J15 and 18 from jalebi batter) under acidic pH conditions aValues are mean ± SD of three experiments |
 | Table 2- Survival rate of the isolates (Id 9, 11, 14, 15 and 18 from idli batter; J15 and 18 from jalebi batter) at different oxbile concentrations and at different time intervals a Values are mean ± SD of three experiments |
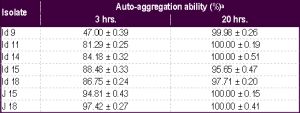 | Table 3- Auto-aggregation ability of the yeast isolates a Values are mean ± SD of three experiments |
 | Table 4- Cell surface hydrophobicity of the selected yeast isolates a Values are mean ± SD of three experiments |
 | Table 5- Morphological characteristics of yeasts isolated from idli and jalebi batter |