ISSN : 0976-9927
EISSN : 0976-9935

TRAIFALGAR R.F.M.1*, KOSHIO S.2, ISHIKAWA M.3, SERRANO A.E.4, CORRE V.L.5
1Institute of Aquaculture, College of Fisheries and Ocean Sciences, University of the Philippines in the Visayas, Miag-ao, Iloilo, Philippines.
2Laboratory of Aquatic Animal Nutrition, Faculty of Fisheries, Kagoshima University, Kagoshima, Japan.
3Laboratory of Aquatic Animal Nutrition, Faculty of Fisheries, Kagoshima University, Kagoshima, Japan.
4Institute of Aquaculture, College of Fisheries and Ocean Sciences, University of the Philippines in the Visayas, Miag-ao, Iloilo, Philippines.
5Institute of Aquaculture, College of Fisheries and Ocean Sciences, University of the Philippines in the Visayas, Miag-ao, Iloilo, Philippines.
* Corresponding Author : skerferd@yahoo.com
Received : 31-03-2012 Accepted : 10-05-2012 Published : 14-05-2012
Volume : 3 Issue : 1 Pages : 33 - 36
J Fish Aquaculture 3.1 (2012):33-36
DOI : http://dx.doi.org/10.9735/0976-9927.3.1.33-36
Feeding trial was conducted to evaluate the effects of dietary fucoidan supplementation on biological performance and bacterial resistance of early larval stage of Penaeus japonicus. Four microbound semi-purified diets containing graded levels of fucoidan (0.0, 100, 500, 1000 mg/kg diet) were formulated and fed to zoea larval stage of P. japonicus. Each diet was fed to three replicate groups of larvae and the feeding trial was terminated after the larvae had metamorphosed to the mysis stage. Results indicate that supplementation level of 500 mg fucoidan/kg diet is adequate to improve the zoea metamorphic survival to the mysis stage and this supplementation level was also found to be optimum in enhancing the bacterial resistance of zoea larvae against pathogenic Vibrio harveyi infection. The present results suggest that Undaria pinnatifida fucoidan at a dose of 500 mg/kg diet could be used as a dietary supplement to improve the metamorphic survival and enhance bacterial resistance of zoea larval stage of P. japonicas.
Fucoidan, zoea, Penaeus japonicus, Vibriosis resistance.
In the past decades, increasing global demands for shrimp has led to a rapid expansion and intensification of shrimp culture causing serious environmental deteriorations and emergence of infectious agents implicated in severe production losses both in hatchery and in grow-out operations [11] . To date, microbiological diseases are considered as major obstacle in the sustainability and profitability of penaeid shrimp aquaculture. The industry has experienced serious economic losses due to the out breaks of pathogens that include fungi, bacteria and viruses [28] . Heavy mortalities in ponds are usually attributed to virus infections, while in hatcheries the mortalities are mainly caused by fungal and bacterial infections. Several evidences also suggest that the early larval stages of shrimp are more susceptible to bacterial infections than the juvenile and adult stages [7,10,15,18] .
The use of immunostimulating compound has been proposed as a potential means in reducing the risk of production losses due to disease outbreaks in shrimp aquaculture. Most immunostimulants identified to date are compounds derived from cellular components of bacteria and fungi. Enhancement of immune responses leading to the improvement of resistance against infectious agents and better growth performance are the known documented effects of immunostimulants in several species of shrimp [14,19,22,24] . Moreover, polysaccharides of algal origin such as carrageenan, laminaran, alginate and fucoidan have also been shown to exhibit potent immunomodulatory effects on shrimp [29,4,5,16] . However, in contrast to most microbial-derived immunostimulants such as β-glucan, peptidoglycan, and lipopolysaccharide, which effects on shrimp are well-established, information regarding the effects of seaweed polysaccharides on shrimp larval physiology and disease resistance has been relatively scarce.
Fucoidan, a fucose-enriched sulfated polysaccharide from brown algae has been documented in vertebrates to elicit potent antiviral and immune enhancing properties [3] . In addition, dietary supplementation of fucoidan from Cladosiphon okamuranus and Sargassum polycystum has been documented to improve the immunological response and resistance against viral infection in juvenile M. japonicus [25] and P. monodon [5] respectively. Although these results are promising, but the effects of fucoidan on the biological performance and bacterial infection resistance of the early larval stage of shrimp have not been evaluated to date.
The present work was conducted to determine the influence of dietary fucoidan supplementation on the metamorphosis survival and bacterial infection resistance of the early larval stage of Penaeus japonicus.
Fucoidan preparation and test diets
Fucoidan was prepared from a dried sporophyll powder of U. pinnatifida (Marui products Co., Ltd., Kitakyushu, Japan), following the scheme described by Mori and Nisiziwa (1982). The fucoidan polysaccharide has a fucose and sulfate content of 40.0 % and 23.5 % respectively.
Composition of the basal diet [Table-1] used in this feeding trial was patterned based on the formulation used in the larval nutritional experiment with P. japonicus by Koshio et al. (1991). Four dietary treatments were used consisting of increasing dietary inclusion levels of fucoidan (0, 100, 500, 1000 mg/kg diet). The Experimental diets were prepared in microbound form following the procedures described by Teshima et al. (1982). The diets were prepared by mixing the dry ingredients with feed oil followed by the addition fucoidan dissolved in adequate amount of water. The binder zein dissolved in 70% ethanol was then added and mixed to the resulting moist mash. Ethanol was removed from the mixture by evaporation under reduced pressure and the resulting diet were freeze dried, crumbled and sieved to appropriate sizes. Diets were stored at - 28 °C until used.
aHighly unsaturated fatty acids; a mixture of eicosapentaenoic (20:5n-3) and docosahexaenoic acid (22:6n-3) ethyl esters (3:2, w: w; Riken Vitamin Co. Ltd., Tokyo, Japan).
b(mg/100 g): p-amino benzoic acid,9.48; d-Biotin,0.38; Inositol,379.20; Niacin,37.92; Ca-pantothenate, 56.88; Pyridoxine-HCl, 11.38; Riboflavin, 7.58; Thiamin-HCl,3.79; l-ascorbyl-2-phosphate-Mg,132.00; Folic acid, 0.76; Cyanocobalamine, 0.08; Menadione, 3.80; Vitamin A-palmitate,17.85; α-tocopherol,18.96; Calciferol, 1.14.
c(g/100Â gdiet): K2PO4, 2.011; Ca(H2PO4)2.2H2O, 2.736; MgSO4.7H2O, 3.05; NaH2PO4.2H2, 0.795.
The larvae used in this study were obtained from wild spawners caught from Miyasaki prefecture, Japan. The trial was run using zoea stage larvae (Z1). Groups of 100 Z1 were randomly stocked into 12 1-L beakers containing 750 ml of 34 ppt, membrane filtered (50µm) and UV sterilized seawater. Aeration for each beaker was supplied through a glass Pasteur pipette with tip reaching to the bottom, providing a mild stream of air that circulated water inside the container, keeping feed particles in suspension. These culture containers were suspended in a 28oC water bath and were maintained under normal photoperiod. Each experimental diet was fed to triplicate groups of Z1 until reaching complete metamorphosis to the Mysis stage (M1). Treatment groups were fed their respective diets (microbound diet, 53-100 µm) at a rate of 0.16 mg/larvae, subdivided into two equal feedings at 0900 and 1700 h. Quantification of survival, larval stage identification and complete renewal of rearing water was conducted daily following the scheme described by Koshio et al. (1991). Larval stages were determined as described by Hudinaga (1942).
Another group of larvae were maintained with the test diets and exposed to pathogenic Vibrio harveyi via immersion to asses the efficacy of dietary fucoidan supplementation in enhancing the resistance of zoea against vibrio infection. The bacterial pathogen used is a virulent Vibrio harveyi strain (PN-9801), obtained from the bacterial collection of SEAFDEC Aquaculture Department, Philippines. This bacterial strain has been maintained and cultured in thiosulfate citrate bile salt sucrose agar media (TCBS, Difco) and tryptose soy broth (TSB, BBL) supplemented with 2% NaCl. Five treatment groups consisting of 300 Z1 larvae per treatment were reared and fed with the experimental diets containing graded levels of dietary fucoidan (0, 100, 500, 1000 mg/kg) until the larvae has molted to Zoea2 (Z2) stage. From each treatment, 200 Z2 larvae were collected and stocked to 1L round bottom glass containers filled with 750 ml of autoclaved natural seawater and aerated through 0.22µm filters. Immersion pathogen challenge test was then conducted in accordance to the method described by Praytino & Latchford (1995) and Soto-Rodrigues et al. (2006). Larvae were infected by immersion in an aerated viable suspension of V. harveyi (PN-9801)at a concentration of 107 CFU/ml. A separate group of larvae not exposed to the bacterial pathogen served as the non-infected control group. The larvae were not fed during the duration of the test and surviving larvae were counted after 48 h.
Mean metamorphic larval survival among the treatments were compared using one-way Analysis of Variance (ANOVA) and significant differences among the treatments were resolved using Tukey’s honest significant difference test (super ANOVA, ver.1.11, Abacus Concepts, Berkeley, California, USA) at 0.05 level of significance. Chi square test was used to compare the differences in survival response among the treatments following the bacterial challenge test.
Metamorphosis survival of the zoea larvae to the mysis stage (M1) as a response to dietary fucoidan supplementation is shown in [Fig-1] . Significantly higher metamorphic survival were observed in treatments fed diets supplemented with 500 and 1000 mg fucoidan as compared to treatments fed the control and the lowest fucoidan supplementation. The difference in survival in treatments receiving 500 and 1000 mg fucoidan supplementation was not significant. Metamorphic survival in treatment receiving100 mg fucoidan supplementation did not differ with the control treatment and was the lowest among the dietary treatments.
Dietary fucoidan supplementation was observed to improve the resistance of Zoea2 (Z2) larvae against the pathogenic bacteria, V. harveyi [Fig-2] . Mean post-challenge survival of larvae increased with increasing level of dietary fucoidan supplementation. Highest survival was observed in dietary treatment with 1000 mg fucoidan supplementation followed by the treatment with 500 mg supplementation. No significant difference in the number of surviving larvae was observed in these treatments. The control treatment exhibited the lowest survival that was not significantly different to that of treatment with 100 mg fucoidan supplementation. Larval survival in these dietary treatments was significantly lowr
than that of treatments with 500 and 1000 mg fucoidan supplementation.
In recent years, there have been a considerable number of works highlighting the potential benefits of immunostimulants to enhance the immunocompetence and disease resistance of cultured shrimp. However, these earlier studies were centered on understanding the modulation of immunological responses related to disease resistance in juvenile and sub-adult shrimp and concerns regarding the influence of immunostimulants on larvae have not been fully addressed. Crustacean larva has a large body surface area and undergoes radical changes both in morphology and physiology during the course of development, making it highly vulnerable to microbial infections and environmental stressors [21] . Immunostimulation has been suggested as a potential method for improving survival by enhancing the larval immunocompetence against pathogenic agents [27] . However, the current knowledge on the biological response and physiological consequences of immune stimulation in larvae has been limited. Thus, the present study was conducted as the first attempt to evaluate the influence of dietary administered immunostimulant (fucoidan) on the success of metamorphosis and vibriosis resistance of P. japonicus zoea larvae.
The results for larval metamorphic survival achieved in the present study are comparable with previous reports in shrimp larvae reared with experimental diets under laboratory conditions [9,1,6] , indicating that the dietary formulation containing fucoidan is acceptable and not detrimental to the larval physiological processes. Relative to the control, significant enhancement of metamorphic survival from zoea - mysis is evident in groups receiving 500 mg / kg supplementation with higher supplementation level at 1000 mg/ kg provided no additional improvement in survival. This suggests that supplementation level of 500 mg/kg is adequate to enhance larval metamorphosis from zoea to the mysis stage. Similar to the present findings, it was also documented that fucoidan administered by immersion accelerates the embryonic development and increase the life span of developing sea urchin embryos suggesting a cellular growth promoting activity of this compound [12] . Morover, Itami et al., (1991) reported that dietary supplementation of a vibrio bacterin as an immunostimulant improved the survival of P. monodon larvae. Improved larval survival was also reported in H. hippoglosus larvae immunostimulated by feeding with artemia-bioencapsulated alginate [20] and by immersion in A. salmonicida lipopolysaccharide. The present results elucidating the enhancement of larval survival by fucoidan supplementation is the first time to be documented in a penaeid shrimp and concurs with these previous findings, emphasizing the benefits of dietary immunostimulant in improving the larval metamorphic survival.
In juvenile shrimp, enhancement of growth performance and improvement of health conditions are commonly observed as positive benefits of immunostimulant supplementation [9,22] . These effects have been hypothesized as a consequence of enhanced nutrient digestion and assimilation caused by the activation of fixed hemocytes that secrete hydrolytic enzymes in the digestive gland [2] . Moreover, immunostimulants including fucoidan are known to enhance hematopoiesis in shrimp [25,5] . Hemocyte functions as a major organ for immunological defense, for wound healing and for nutrient storage and transport. The improvement of larval survival observed in the present study could be a result of enhanced hemocytic activity similar to that observed in juvenile shrimp. Enhancement of metamorphic survival could also be attributed to efficient utilization of cellular energy reserves. In rats, it was shown that feeding with diet supplemented with Undaria pinnatifida, (the source of fucoidan in this study) enhanced the utilization of lipids caused by the activation of enzymes involved in lipid metabolism [3,17] . Similar effects on efficient utilization of energy reserves resulting in improved health and biological performance has also been reported in red sea bream fed diets supplemented with Ascophylum nodusom and Undaria pinnatifida algal meals [30] .
Fucoidan has been known as a potent activator of cellular immune response in terrestrial vertebrates [3] . Similarly, the current study demonstrates that dietary fucoidan supplementation can also enhance the resistance of shrimp zoea larvae against V. harveyi infection. Survival percentage increased with increasing fucoidan concentration in the diet. As compared to the control, higher survival is evident at 100 mg/kg supplementation with optimum effects noted at 500 mg/kg supplementation level. Previously it was also reported that dietary fucoidan supplementation enhanced the resistance of juvenile M. japonicus [25] and P. monodon [5] against white spot syndrome virus infection. The authors attributed these protective effects to the enhancement of hemocyte phagocytic activity and to the inhibition of viral adsorption to the host. The present results provide additional evidence that dietary fucoidan supplementation could also promote larval resistance against bacterial infection.
Although the larval immune physiology is not yet fully understood, recent evidence suggests that shrimp larvae are capable of producing antibacterial peptides in their hemocytes as a response to the presence of infectious and immune stimulating agents. In addition, it has been suggested that the mode by which fucoidan enhances shrimp immune response is by the activation of a phagocytosis activating protein, known to initiate and enhance hemocyte phagocytic activity [5] . Although the hematological and immunological response was not measured in this study, it is conceivable that the enhancement of survival after the challenge may have been due to the activation of cellular immune response, similar to that observe in juvenile shrimp.
Overall the present findings indicate that dietary fucoidan supplementation at a dose of 500 mg/kg diet could improve the metamorphosis survival and enhance the resistance of zoea larvae against pathogenic V. harveyi infection. Moreover, fucoidan supplementation could be used as prophylactic means to improve hatchery production of P. japonicus.
The scholarship support by the Ministry of Education, Culture and Sports of Japan for the realization of this work is highly acknowledged.
[1] Alabi A.O., Cob C., Jones D.A., Latchford J.W. (1999) Aquaculture Int., 7, 137-158.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[2] Azad I.S., Panigrahi A., Gopal C., Paulpandi S., Paulpandi S., Mahima C., Ravichandran P. (2005) Aquaculture, 248, 227-234.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[3] Berteau O., Mulloy B. (2003) Glycobiology, 13, 29-40.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[4] Cheng W., Liu C.H., Kuo C.M. Chen J.C. (2005) Fish Shellfish Immunol.,18, 1-12.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[5] Chotigeat Tongsupa W.S., Supamataya K., Phongdara A. (2004) Aquaculture, 233, 23-30.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[6] D′Abramo L., Perez E.I., Sangha R., Puello-Cruz A. (2006) Aquaculture, 261, 1356-1362.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[7] Hameed A.S. (1995) J. World Aquaculture. Soc., 256, 315-319.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[8] Hudinaga M. (1942) Jpn. J. Nut.,111, 425-431.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[9] Itami T., Takahashi Y., Yoneoka K., Yan Y. (1991). J. Aqua. Ani. Health 3, 151-152.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[10] Jiravanichpaisal P., Narongsak P., Sasithon P., Donnuea S., Söderhäll I., Söderhäll K. (2007). Fish & Shellfish Immunol., 23, 815-824.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[11] Karunasagar I., Pai, R., Malathi G.R., Karunasagar I. (1994) Aquaculture, 128, 203-209.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[12] Kiseleva M.I., Shevchenko N.M., Krupnova T.N., Zvyagintseva T.N. (2005) J.Evo. Biochem. Physiol., 41, 51-58.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[13] Koshio S., Kanazawa A., Teshima S. (1991) Nip. Sui. Gak., 58, 1083-1089.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[14] Koshio S., Yokoyama S., Ishikawa M., Hayasawa H., Oshida K. (2000) Lactoferrin Structure, functions and applications, 451-455.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[15] Lavilla-Pitogo C.R., Baticados M.C.L., Cruz-Lacierda E.R., De La Pena L.D. (1990) Aquaculture, 91, 1-13.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[16] Martin G., Poole D., Poole C., Hose J.E., Arias M., Reynolds L., McKrell N., Whang A. (1993) J. Invert. Pathol., 62, 308-315.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[17] Mori H., Nisiziwa K. (1982) Bull. Jpn. Soc. Sci. Fish. 48, 981-986.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[18] Prayitno S.B., Latchford J.W. (1995) Aquaculture, 132, 105-11.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[19] RodrÃguez J., Espinosa Y., EcheverrÃa F., Cárdenas G., Román R., Ster S. (2007) Aquaculture, 273, 405-415.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[20] Skjermo J., Bergh Ø. (2004) Aquaculture, 238, 107-113.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[21] Smith V.J., Brown J.H., Hauton C. (2003) Fish Shellfish Immunol. 15, 71-90.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[22] Song Y.L., Sung H.H. (1990) Bull. Eur. Assoc. Fish Pathol., 10, 4-98.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[23] Soto-Rodriguez S.A., Simoes N., Roque A., Gomez Gil B. (2006) Aquaculture, 109-115.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[24] Sung H.H., Kuo G.H., Song Y.L. (1994) Fish Pathol., 29, 11-17.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[25] Takahashi Y., Uehara K., Watanabe R., Okumura T., Yamashita T., Omura H., Yomo T., Kanemitsu A., Kawano T., Narasaka H., Suzuki N., Itami T. (1998) Advances in Shrimp Biotechnology, Published by the National Center for Genetic Engineering and Biotechnology, Bangkok, Thailand, 171-173.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[26] Teshima S., Ishikawa M., Alam M.D. Koshio S., Michael F.R. (2004) Comp. Biochem. Physiol., Part B 137, 209-217.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[27] Vadstein O. (1997) Aquaculture 155, 401-417.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[28] Vaseeharan B., Ramasamy P. (2003) Microbiol. Res., 158, 299-308.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[29] Yeh S.T., Chen J.C. (2008) Aquaculture, 276, 22-28.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
[30] Yone Y., Furuichi M., Urano K. (1986) Bul. Japn. Soc. Sci. Fish., 52, 1465-1488.
» CrossRef » Google Scholar » PubMed » DOAJ » CAS » Scopus
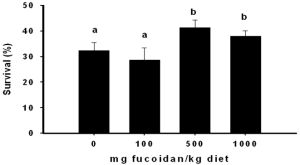 | Fig. 1- Metamorphosis survival to mysis stage of zoea larvae fed fucoidan supplemented diets. Bars are means of triplicate groups ± S.E. Mean values bearing similar superscript are not significantly different. |
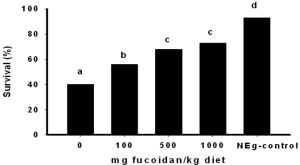 | Fig. 2- Survival of zoea larvae fed fucoidan supplemented diets and challenged with pathogenic Vibrio harveyi by immersion. Values bearing similar superscript are not significantly different (χ2 -test,α= 0.05). |
 | Table 1- Composition of the basal diet |